
Polychaetes, segmented worms bearing parapodia, are first found in the Cambrian Period, and all exhibit an elongated body broken into many segments. More specifically, the Polychaete worms within the order Eunicida are also first found in the Cambrian and are further characterized by ventral maxillae and dorsal mandibles positioned within the pharynx. Eunicids are found in many ocean environments today, with 443 species in the family Eunicidae alone (comprising half of all known modern species), but have a rich fossil record which is twice as diverse as those still alive today (1).
Polychaetes found at Seven Stars
Eunicida indet.
UNCOMMON
How do I identify Polychaetes?
Because of their soft, unmineralized bodies, fossil Eunicids are primarily known from their oral apparatus, composed of numerous sclerotized plates called scolecodonts (4). At first glance, they may appear similar to Conodont elements, but on closer inspection they exhibit obvious differences. They are composed of a calcium carbonate cuticle (which can be molted similarly to arthropod exoskeletons in some Paleozoic Eunicids and modern Labidognaths) (4) and are used for scraping, capturing prey, and shredding, depending on the ecological niche occupied by the Eunicid (1).
Unfortunately, no scolecodonts have been recovered from Seven Stars. The only clue of Eunicid presence at the site is the existence of rounded, striated burrows that are found in grey shale at the upper collection area.
Scolecodonts have been found in other Hamilton group formations, and have been assigned to the wastebasket taxon, Arabellites, which was used for individual teeth that looked similar to those of a modern genus (12). The Hamilton scolecodonts are strikingly larger than many other Paleozoic scolecodonts, and exhibit novel structure (13).
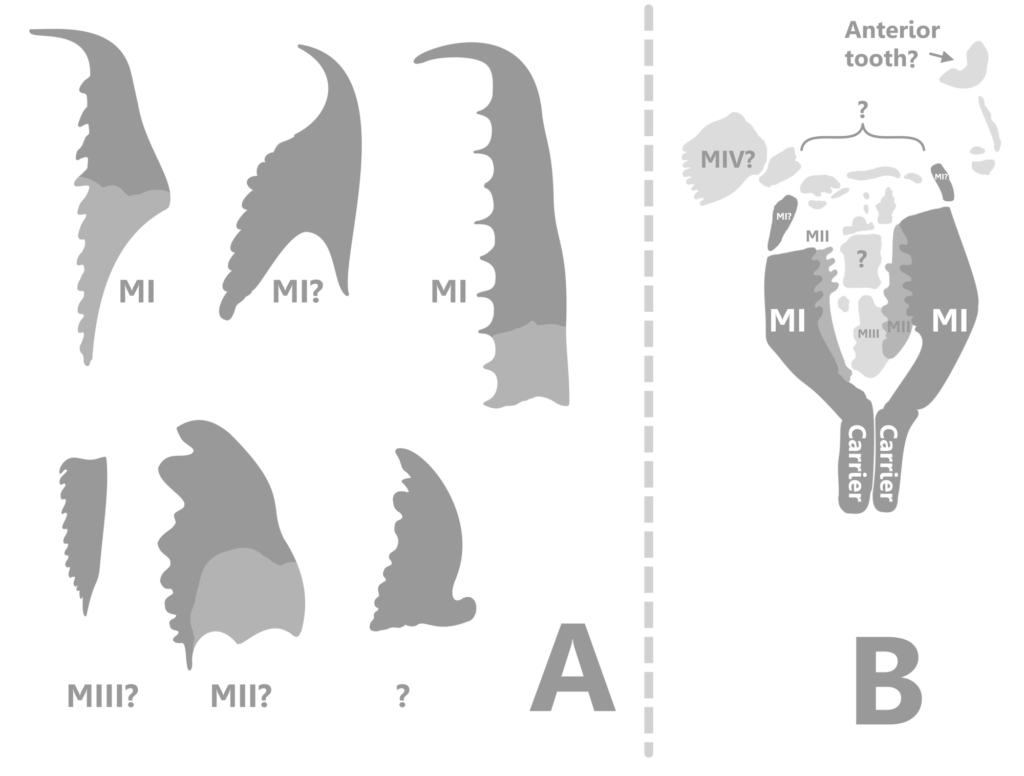
The Hamilton scolecodonts appear similar in form to modern Labidognaths, and may have been shed periodically because of wear and continued growth (7, 8, 10). There is no evidence that the Polychaetes that lived at Seven Stars are the same species that are found in other Hamilton sites, which lived in different depositional environments. However, there is no positive evidence of any other organisms that would dig burrows similar to those that have been found.
The Polychaete burrows can be differentiated from fossilized branches washed in from rivers because of the repeated scrape marks made by the burrowing worm within. Their paths also undulate and meander unlike the mostly straight Archaeopteris fossils that are sometimes found in the underlying strata. The worm burrows cut through multiple strata more rarely than Archaeopteris branches, and when they do, they are less circular in cross section and have a crystal internal structure instead of a pyritized hollow core.
What are Polychaetes?
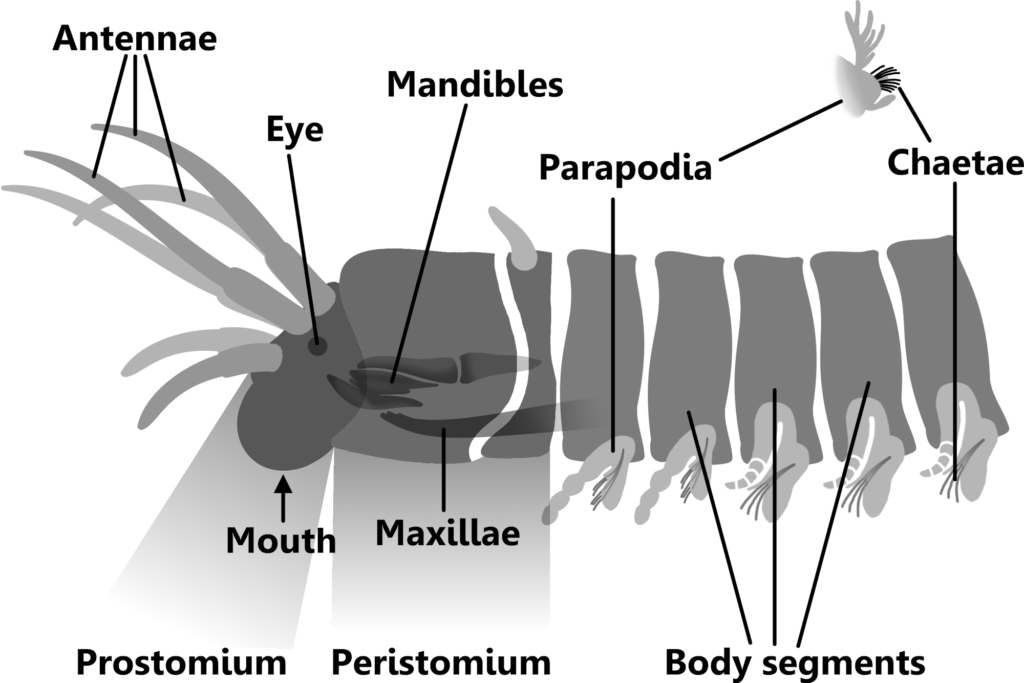
Polychaetes, or bristle worms, are all characterized by segmented bodies, each segment bearing a pair of parapodia. These are projections that bear chaetae (5), which will be discussed below.
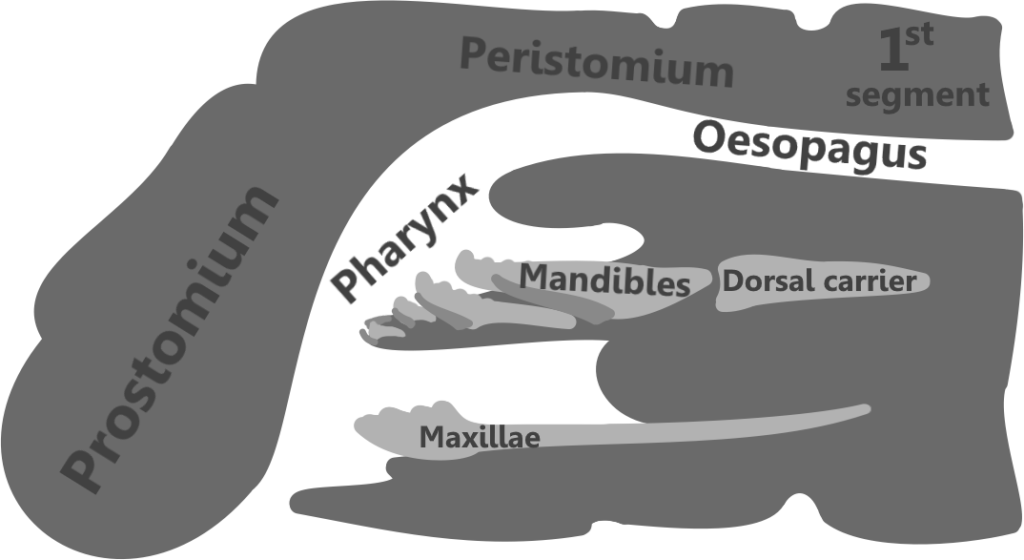
The first segment of the body is called the prostomium, which bears eyes and sensory antennae. The presence and configuration of these and other structures vary within different families (5), but Eunicids usually have antennae and sometimes eyes (1).
The second segment is called the peristomium, which bears the mouth and oral apparatus. The pharynx, the mouth and esophagus, is eversible, being able to be turned inside out for feeding and burrowing purposes (5). In Eunicids it is ventral, with the maxillae and mandibles positioned below it (6).
After the Prostomium and Peristomium, the body is broken into many segments, each having a pair of parapodia. These were ventral or lateral projections that support the chaetae (tufts of spines used for locomotion), a leg-like protrusion, and in some species, feathery gills. The parapodia can be used for walking, swimming, food manipulation, and aid in burrowing (5).
While many polychaetes have jaws, the Eunicids are most notable for their complex, diverse apparatuses that fossilize well and have a wide range of shapes, orientations, and functions. Modern Eunicid jaws can be used to differentiate species, because of their great diversity (1).
Fossil Eunicid jaws are called scolecodonts, which literally translates from Latin meaning “worm jaws”. They are found from late Cambrian strata to the present, and seem to have been important members of the Paleozoic marine fauna (4). Throughout the 20th century, Paleontologists used their own nomenclature for the scolecodonts they found, referring to their structure and often missing the point that each individual worm’s oral apparatus was composed of many different, and often asymmetrical, individual jaws. As with Conodonts, each new form encountered was given its own genus and species, instead of the sometimes very difficult task of piecing together the entire apparatus from assemblages and knowledge of different groups and their characteristics. This led to many, many species being erected for the same species of worm (4, 13 – example).
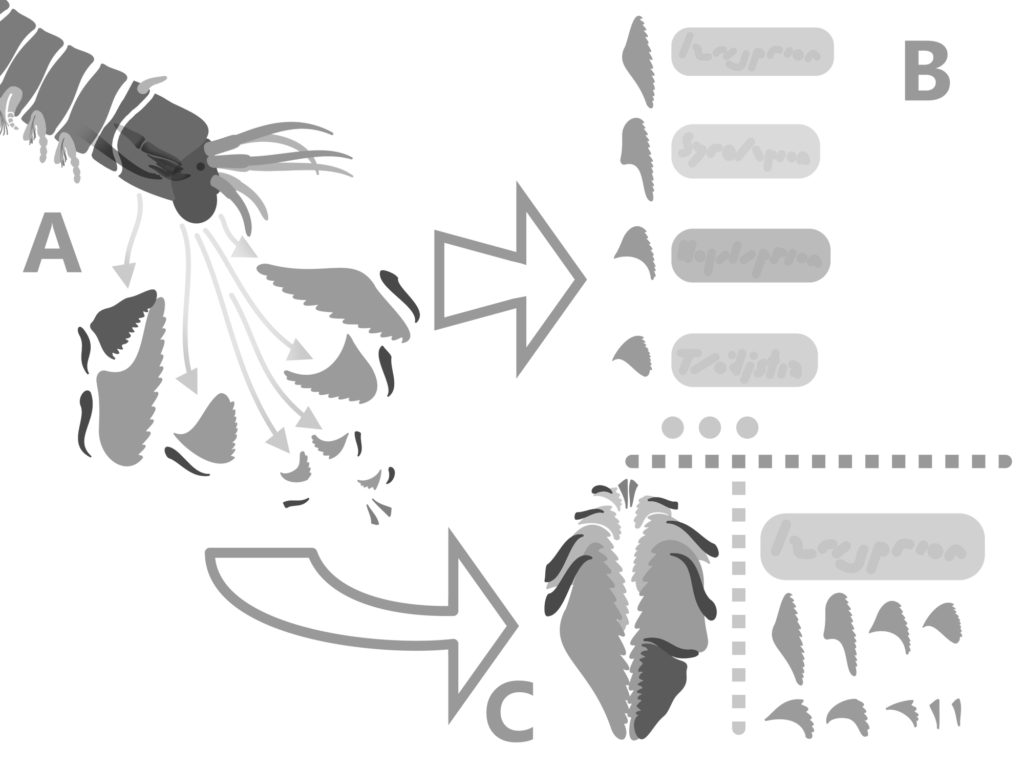
Once scolecodonts have been organized into entire apparatuses, their interrelationships can be put together (15). Overall form of elements, as well as their size and placement, is helpful in producing separate groups (2). This is the morphologic path of classification within Eunicida; it is the only possible way to include fossil forms (2), but often conflicts with more recent molecular studies using selected genes from modern groups. Because these molecular studies often have conflicting results, and because of the fact that they cannot be used to classify fossil taxa, the morphological strategy of classification is favored here as a more accurate method.
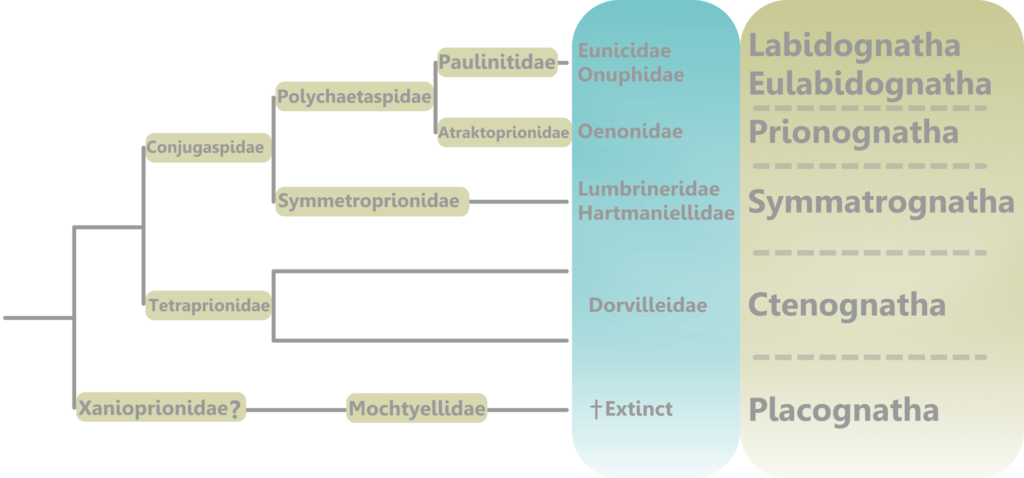
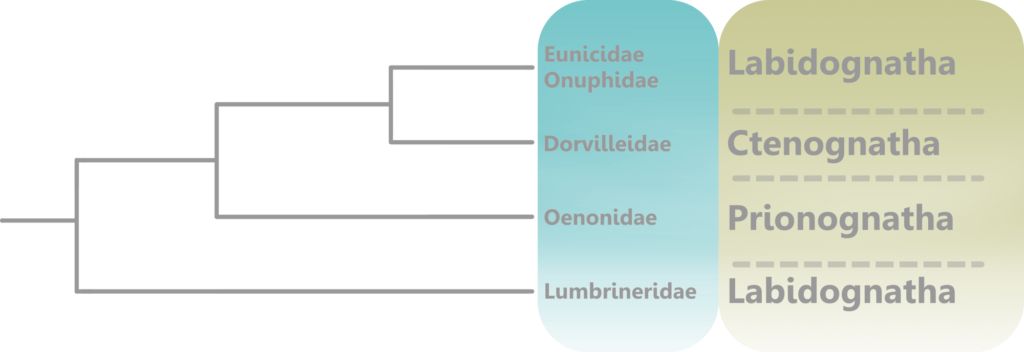
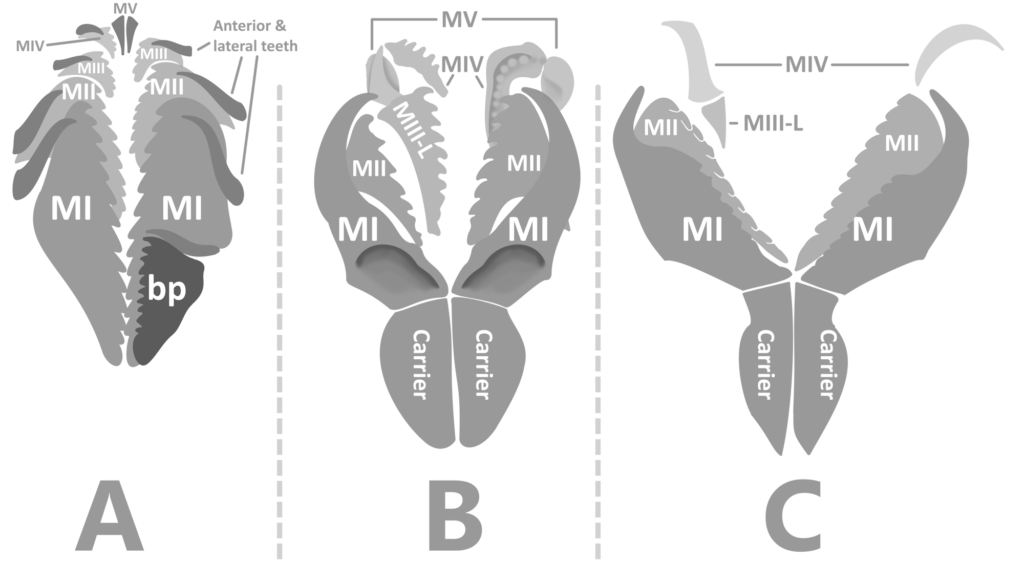
In the living animal, the jaws would have been posterior to the mouth, and under the esophagus (6). They were eversible, meaning that they could be extended out of the pharynx and used for burrowing, capturing prey, processing food, and more (5). They could then be retracted back into the pharynx when the worm was not feeding or taking part in such activities, for protection, continued growth, and convenience.
In Labidognath Eunicids, the jaws were molted similarly to arthropod exoskeletons, where the old jaw pulls away from the secreting tissues (apolysis) of the mouth, and is released (ecdysis). This reveals a larger (x1.4) jaw beneath it, which is white before sclerotization in modern labidognaths (8). Fossils from the Ordovician period reveal the imprint of a new jaw in the pulp cavity of the old jaw, providing direct evidence that Paleozoic Eunicids molted their jaws as modern examples do (7, 10).
Eunicids build their jaws from calcium carbonate, but harden the tips with zinc, iron, or iron and titanium, depending on the worm in question (9). If the jaw did break, one study suggests that they could regenerate the break like in Conodonts, but from the inside, instead of the outside (10), until the new jaw underneath was exposed through ecdysis.
Polychaetes at Seven Stars
While their fossil material is not as extensive as other organisms at the site, Eunicids were a regular component of the upper layers of the main collecting area at Seven Stars and can be found alongside the occasional Bivalve or even straight-shelled Cephalopod. As some of the only ichnofossils, they reveal the behavior more than the morphology of the worms that formed them.
Eunicida indet.
The burrows of likely Eunicid worms are sometimes found in the grey boulders and piles on top of the main collecting area. They have faint, repeated grooves on their surface, and their cross section is either homogeneous with the surrounding rock or is crystalized. The burrows are usually about a centimeter in width and wind around on and into the rock surface. No plants are found in these grey rocks, so there is no overlap between Eunicid burrows and similar-looking Archaeopteris branches that are sometimes found in the lower strata.
In the picture to the right, there is a Polychaete burrow lit from the side to show its texture (top) and the same burrow lit from the top to show what it would look like in the field.

Works cited
(1) Zanol, Joana & Carrera-Parra, Luis & Steiner, Tatiana & Amaral, Antonia & Wiklund, Helena & Ravara, Ascensão & Budaeva, Nataliya. (2021). The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity. 13. 74. 10.3390/d13020074. https://www.researchgate.net/publication/349242761_The_Current_State_of_Eunicida_Annelida_Systematics_and_Biodiversity
(2) Paxton, Hannelore. (2009). Phylogeny of Eunicida (Annelida) based on morphology of jaws. Zoosymposia. 2. 241-264. 10.11646/zoosymposia.2.1.18. https://www.mapress.com/zoosymposia/content/2009/v2/f/v002p241-264.pdf
(3) Struck, Torsten & Purschke, Günter & Halanych, Ken. (2006). Phylogeny of Eunicida (Annelida) and Exploring Data Congruence Using a Partition Addition Bootstrap Alteration (PABA) Approach. Systematic biology. 55. 1-20. 10.1080/10635150500354910. https://www.researchgate.net/publication/7269322_Phylogeny_of_Eunicida_Annelida_and_Exploring_Data_Congruence_Using_a_Partition_Addition_Bootstrap_Alteration_PABA_Approach
(4) Olle Hints. (2000-2023). Scolecodonts, the Jaws of Polychaete Annelids. Taltek Department of Geology, Accessed 25 Jan. 2024. https://scolecodonts.net/
(5) Verdonschot, Piet. (2015). Introduction to Annelida and the Class Polychaeta. 10.1016/B978-0-12-385026-3.00020-6. https://www.researchgate.net/publication/285433306_Introduction_to_Annelida_and_the_Class_Polychaeta
(6) Tzetlin, A., Budaeva, N., Vortsepneva, E., & Helm, C. (2020). New insights into the morphology and evolution of the ventral pharynx and jaws in Histriobdellidae (Eunicida, Annelida). Zoological letters, 6(1), 14. https://doi.org/10.1186/s40851-020-00168-2 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7678154/
(7) Mierzejewski, Piotr. (1977). MOLTING of the JAWS of the EARLY PALEOZOIC EUNICIDA (ANNELIDA, POLYCHAETA). ACTA PALAEONTOLOGICA POLONICA, vol. 23. Accessed 25 Jan. 2024. https://www.app.pan.pl/archive/published/app23/app23-073.pdf
(8) Paxton, Hannelore. (2006). El reemplazo de mandíbulas en Eunicida. Scientia Marina. 70. 10.3989/scimar.2006.70s3331. https://www.researchgate.net/publication/250304496_El_reemplazo_de_mandibulas_en_Eunicida
(9) Elfman, Mikael & Eriksson, Mats & Kristiansson, Per & Malmqvist, Klas. (1999). Application of microPIXE and STIM in analyses of fossil and Recent polychaete jaws (scolecodonts). Nuclear Instruments and Methods in Physics Research Section B: Beam Interactions with Materials and Atoms. 158. 287-291. 10.1016/S0168-583X(99)00316-X. https://www.researchgate.net/publication/223003286_Application_of_microPIXE_and_STIM_in_analyses_of_fossil_and_Recent_polychaete_jaws_scolecodonts
(10) Kulashova, Tatiana. (2019). Microornamentation and Regeneration: an Example from Devonian Scolecodonts from the Voronezh Anteclise. Paleontological Journal. 53. 972-976. 10.1134/S0031030119090077. https://www.researchgate.net/publication/339632381_Microornamentation_and_Regeneration_an_Example_from_Devonian_Scolecodonts_from_the_Voronezh_Anteclise
(11) Paxton, Hannelore & Eriksson, Mats. (2012). Ghosts from the past – ancestral features reflected in the jaw ontogeny of the polychaetous annelids Marphysa fauchaldi (Eunicidae) and Diopatra aciculata (Onuphidae). GFF. 134. 309-316. 10.1080/11035897.2012.752762. https://www.researchgate.net/publication/271754480_Ghosts_from_the_past_-_ancestral_features_reflected_in_the_jaw_ontogeny_of_the_polychaetous_annelids_Marphysa_fauchaldi_Eunicidae_and_Diopatra_aciculata_Onuphidae
(12) Eriksson, Mats. (2008). Review of Scolecodonts Assigned to Arabellites, Based on Hinde’s (1879) Type Material. Journal of Paleontology – J PALEONTOL. 82. 628-633. 10.1666/07-090.1. https://www.researchgate.net/publication/250070802_Review_of_Scolecodonts_Assigned_to_Arabellites_Based_on_Hinde%27s_1879_Type_Material
(13) Eller, E. R. (1934). Annelid Jaws from the Hamilton Group of Ontario County, New York. Annals of the Carnegie Museum, vol. 24, 3 Dec. 1934, pp. 51–56, https://doi.org/10.5962/p.215192. Accessed 23 Nov. 2022. https://www.biodiversitylibrary.org/partpdf/215192
(14) Wilson, K. A. (2014). Field guide to the Devonian fossils of New York (3rd ed.). Paleontological Research Institution.
(15) KIELAN-JAWOROWSKA,ZOFIA: Scolecodonts versus jaw apparatuses.
Lethaia, Vol. 1, pp. 39-49. Oslo, 15th January 1968. https://www.idunn.no/doi/pdf/10.1111/j.1502-3931.1968.tb01726.x