
Trilobites were enormously successful arthropods that lived throughout the Paleozoic (2). They are thought to have been most closely related to living horseshoe crabs, but are themselves not chelicerates (3). Four species of trilobites have been found at Seven Stars, but as usual only their hard exoskeletons are preserved.
Trilobites found at Seven Stars

Dipleura dekayi
COMMON
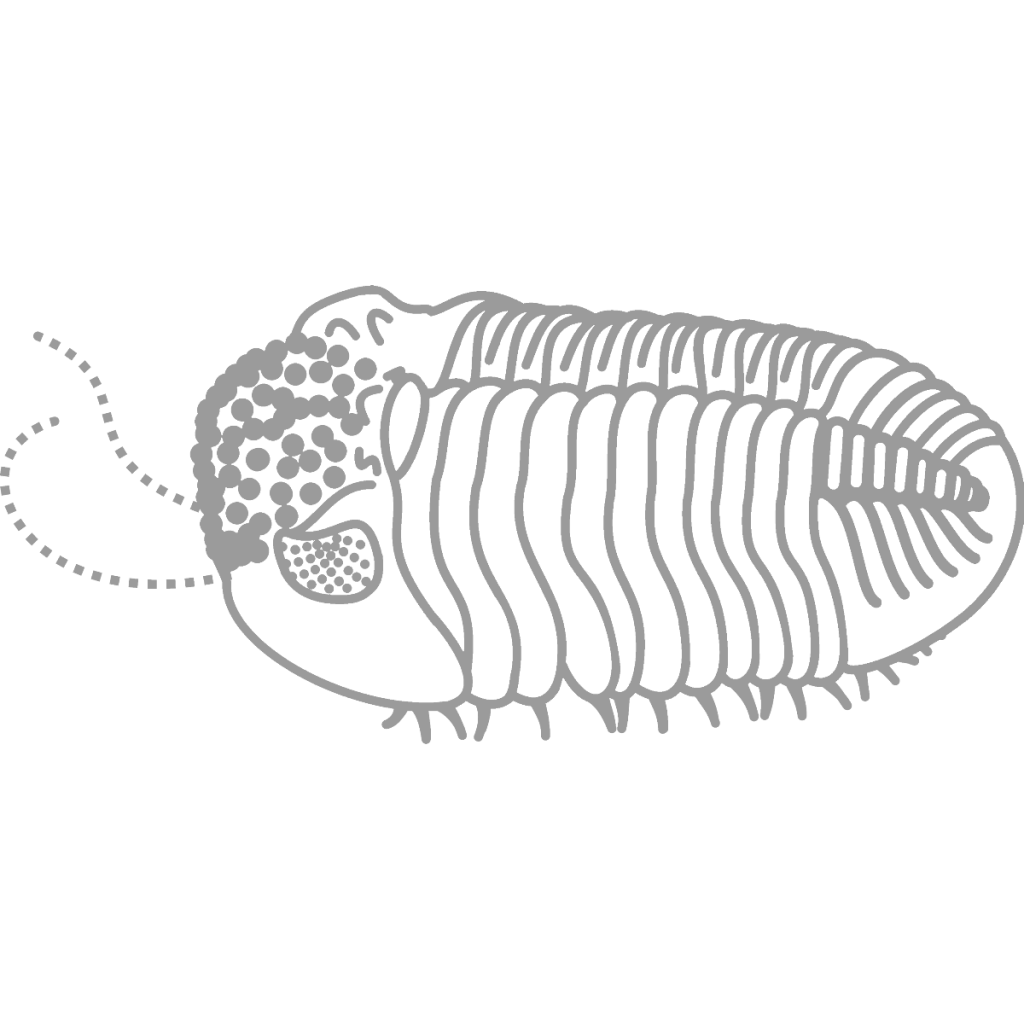
Eldredgeops crassituberculata
VERY RARE

Greenops boothi
RARE
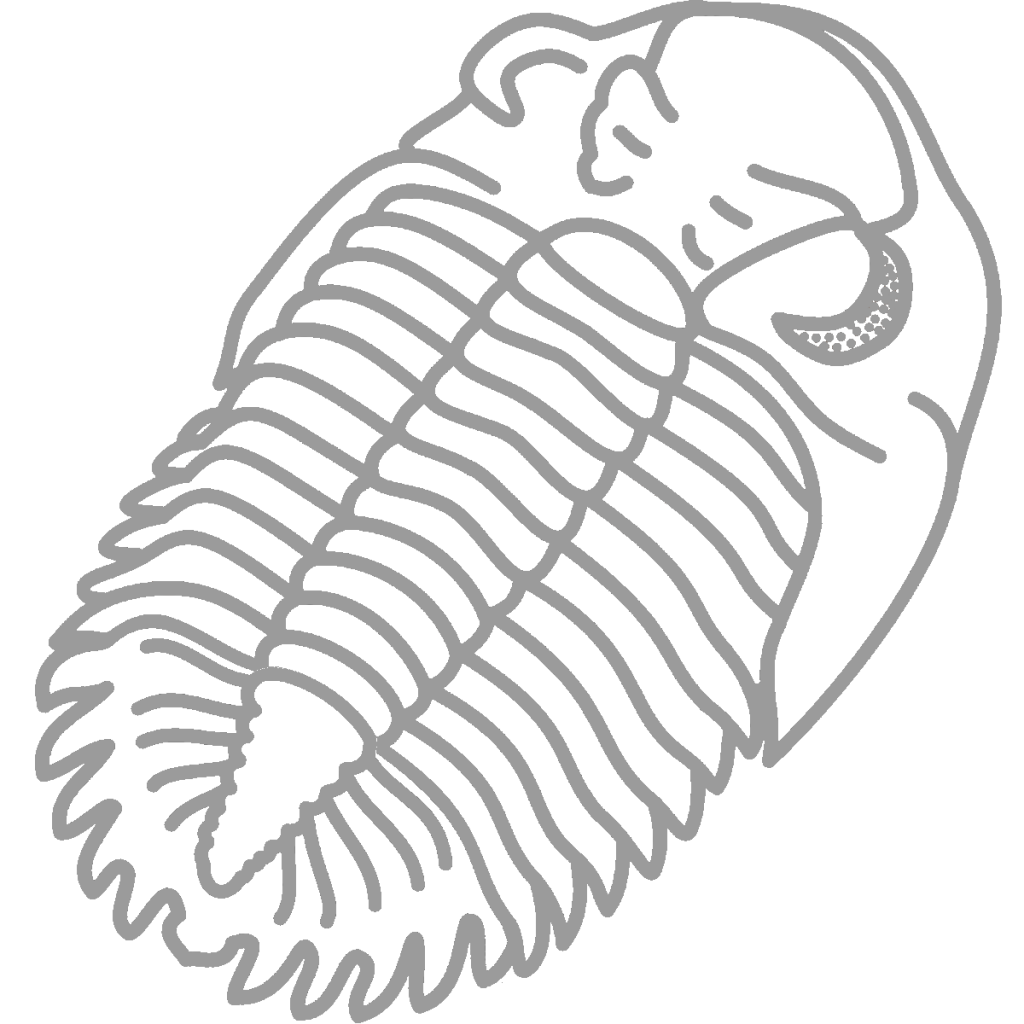
Greenops grabaui
RARE
How Do I Identify Trilobites?
Trilobites are made up of many segments, like all arthropods (1, 2, 3). Different trilobites at Seven Stars look very different as fossils with the exception of the Greenops species. Dipleura can instantly be recognized because of the pores in its shell. The outer layer, in almost all cases, comes off on a different rock and leaves tubular bumps on the fossil (see pictures below). Greenops preserves very differently, its spiny pygidium or tail being the most common find. Eldredgeops is exceedingly rare with only one cephalon or head being found, with a large lump on top of the head.

This Dipleura specimen is instantly recognizable because of its course tubercles.

This Dipleura pygidium (tail) shows further the texture that characterizes Dipleura dekayi.

Greenops and Eldredgeops preserve differently than Dipleura, like this.
What Were Trilobites?
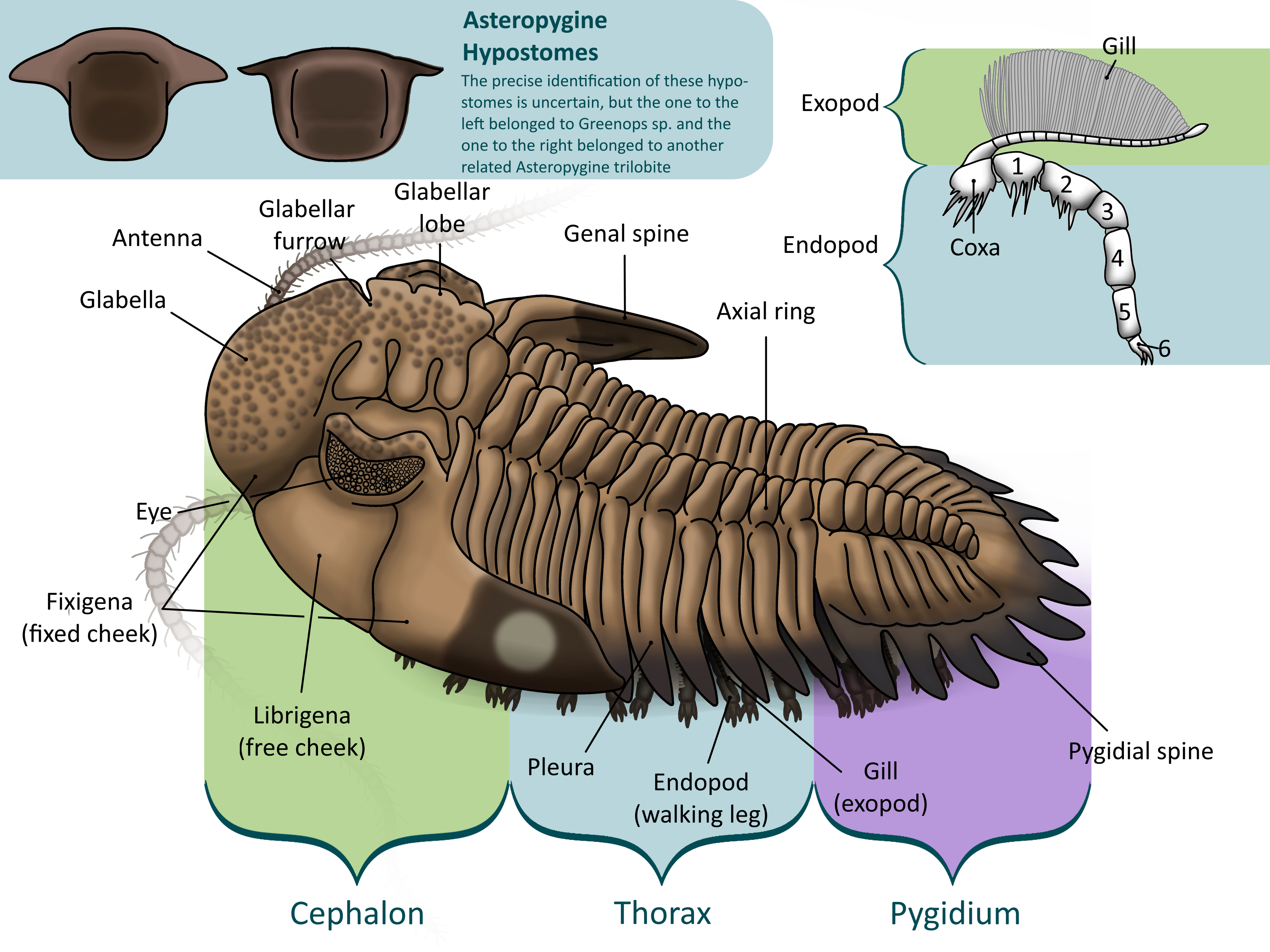
Trilobites blossomed in the Cambrian Period. They were arthropods, like insects, arachnids, and crustaceans, meaning that they had jointed legs and segmented bodies covered in a hard exoskeleton. This was shed periodically to let the trilobite grow (2, 3). Trilobites’ bodies were separated into three main parts – the cephalon, or head, the thorax, or body, and the pygidium, or tail (1, 2, 3).
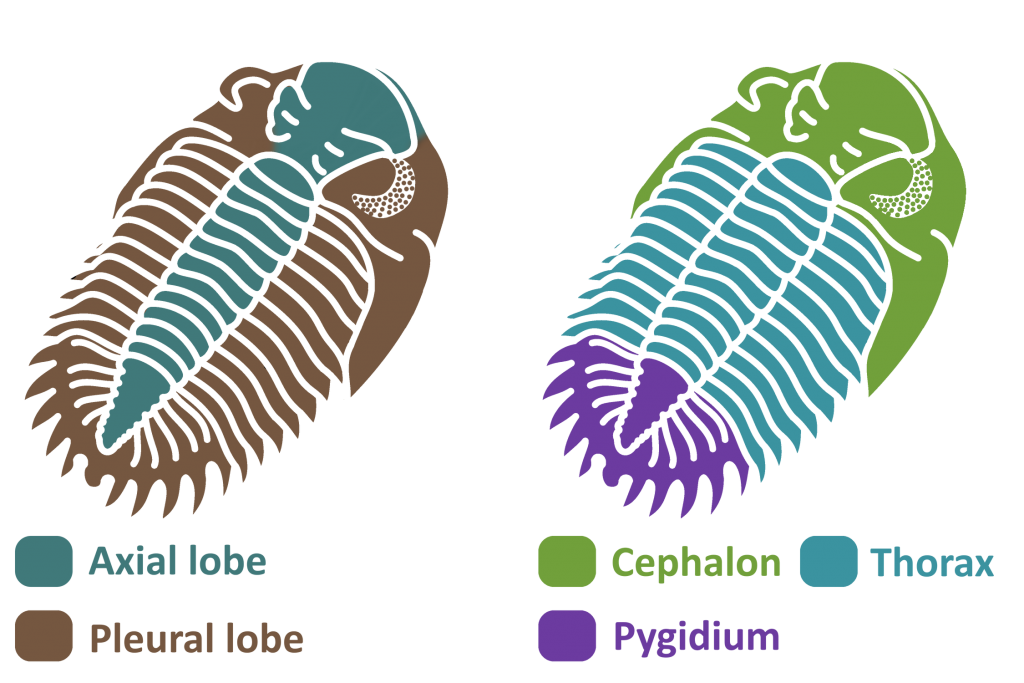
The cephalon was the trilobite’s head. Composed of a couple fused segments, it was covered in plates with edges called sutures. Trilobites had a bulge on the top of their cephalons, called the glabella, that housed the stomach and digestive glands. To the sides of the glabella are the trilobite’s eyes. In most species, these were made up of many lenses, each made of pure calcite (3).
In Phacopids like Greenops and Eldredgeops, each lens was separated from the others by a small ridge, preventing light from one getting into the other (3). This eye type, called the schizochroal eye, allowed for acute vision in low light conditions (4). Eldredgeops had particularly sharp eyesight. Fossils of its very closest relatives reveal that each lens had six more behind it, a phenomenon not seen in any other arthropod (4).
To make the stomach bigger, trilobites had a rounded bulge on the top of the head, called the glabella. The bigger the glabella, the more room there was for the stomach, so it is a general rule that trilobites with larger glabellas preyed on larger organisms in relation to their size than trilobites with smaller glabellas (2).
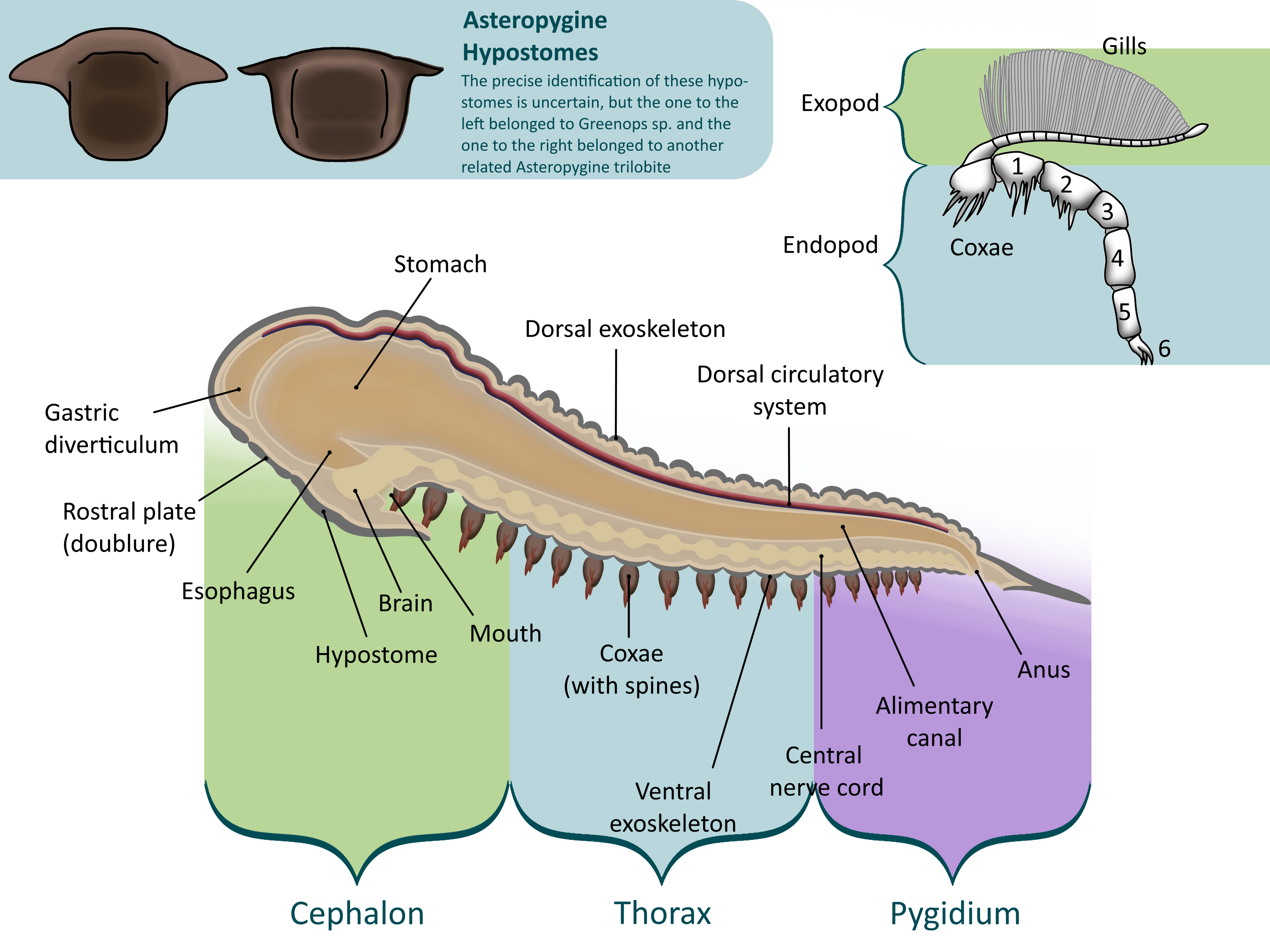
Under the head was a semicircular plate called the hypostome. The hypostome covered the trilobite’s mouth and supported the other organs in the cephalon. In some trilobites, the hypostome was free to move around, called natant. In others, it is fused to a fold in the cephalon called the doublure (conterminant), leading paleontologists to believe that these trilobites were predators. This is because a trilobite could brace prey against a fused hypostome, not as easy with a natant one (2).
The thorax was the trilobite’s body, separated further into many segments. Each segment had three rounded lobes – the central axial lobe and two pleuaral lobes on each side. The axial lobe held the internal organs, muscles, and limbs, while the pleural lobes were simply extensions of the exoskeleton to protect the legs and gills (1, 2, 3).
Each segment held a pair of limbs. Each one had two branches: a walking leg, or endopod, and a second branch, called the exopod, which supported feathery gills (2). This type of limb is called biramous, or two-branched, and was common in early Paleozoic arthropods. The base of the leg, called the coxa (plural coxae), bristled with spines, as did the first few segments of the leg. These were used to shred prey under the trilobite’s body before it was passed up to the hypostome and mouth (2). Trilobite limbs are very rarely preserved, usually in low-oxygen conditions where bacteria cannot survive (3) .
The pygidium was a large plate that covered the last several segments of the trilobite’s body, forming a tail (1, 2, 3) . While the cephalon is made up of fused plates and the thorax is separated into articulated segments, the pygidium was all one piece, making it the most common fossil of the smaller trilobites that tend to break apart. The pygidium held several pairs of legs, as did the thorax and cephalon. The exact number of segments in the pygidium varies from species to species, thus the number of pyginal limbs does too (2).
Most trilobites could roll into a ball when threatened, serving as protection from would-be predators such as cephalopods. In Dipleura and Eldredgeops, the pygidium would be tucked into a vincular furrow under the cephalon, but because of its pyginal spines Greenops tucked its head into its pygidium. Different trilobites enrolled in different fashions. Eldredgeops thrust its head downwards and roll into its protective position under the seafloor where predators like cephalopods would have to dig to get therir prey. It locked its pygidium into its speciallized vincular furrow, with notches to match the pleurae on its pygidium. Greenops was not such an effective enroller, using powerful muscles in its throax/pygidium area to thrust its pygidium into the ground and vault backwards into an enrolled state partially buried in sediment (3). Greenops could not roll as tightly as Eldredgeops could, and compensated with its long spines; these also meant it did not need to grow as thick an exoskeleton, making molting quicker and therefore safer.
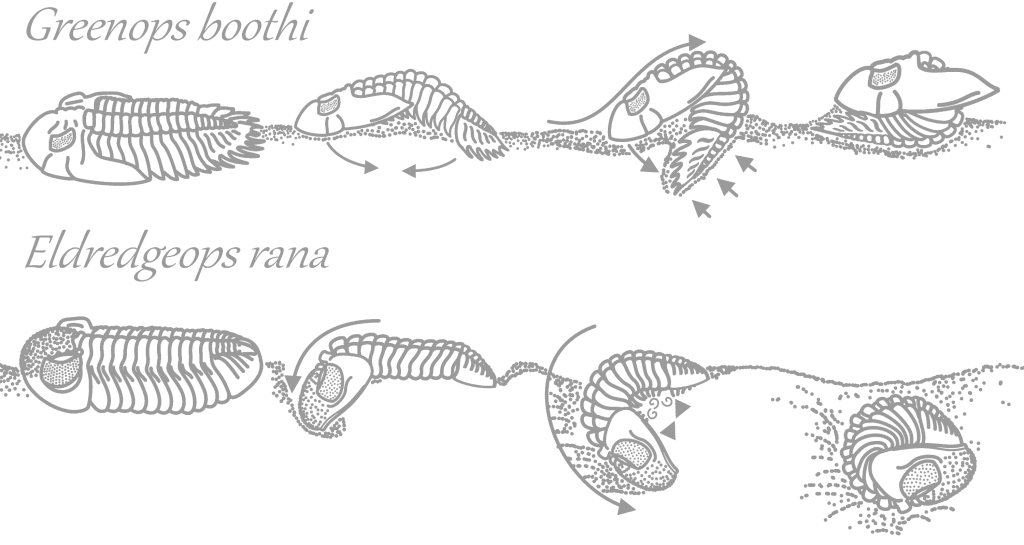
Dipleura dekayi enrolled. The cephalon and pygidium do not lock like Eldredgeops’s do but the exceedingly thick exoskeleton of Dipleura protected it from large cephalopods like Striacoceras.
Greenops grabaui enrolled. The cephalon tucks into a groove in the pygidium but not tightly. Sharp curved spines protect this area and the thorax, where the exoskeleton is most vulnerable to attack.
Eldredgeops crassituberculata enrolled. The pygidium fits tightly into a vincular furrow with notches, under the cephalon. Eldredgeops had strong muscles in its upper thorax and very weak ones in its lower thorax, opposite of Greenops.
Trilobites hatched from eggs roughly a millimeter in size into tiny planktonic swimmers, called the protaspis (2, 3). At first there were no thoracic segments; the cephalon and pygidium were not yet separated. Three pairs of legs poked out from behind a comparatively huge hypostome, propelling the larva and catching other plankton to eat. As the young trilobite grew, its cephalon and pygidium split apart as new segments were added, morphing out of the pygidum, and the trilobite moved to a benthic (living on the seafloor) lifestyle (2, 3).
Like all other euarthropods, molting was required for the trilobite to grow; this process is called ecdysis. Most trilobites molted similarly. Each molt, the librigena, or “free cheeks,” plates on the sides of the trilobite’s cephalon, loosened and fell off, along with the hypostome. The eye lenses are attached to these plates and these were shed along with them. This leaves the glabella and fixigena, or “fixed cheek,” in place (these together are called the cranidium). This plate was less easily removed, as it had to be flipped up so that the trilobite had room to push out of its exoskeleton. Once the fixigena was shed the trilobite was free to walk out of its old thoraxial segments and pyginal plate, quickly grow larger and regenerate injured parts, and grow a new exoskeleton out of calcite. The ventral exoskeleton and limbs were not shed; the same hormone that loosened the librigena softens these areas so that they too can grow larger (3).
Since Phacopidae, the family to which Eldredgeops belonged, did not have functional facial sutures, these trilobites’ librigenas did not separate from the cephalon during ecdysis. Instead, the ventral cuticle of the cephalon became detached from the ventral cuticle of the thorax, and the dorsal cephalic cuticle was pivoted up or rotated to the side, allowing the trilobite to walk out of its exoskeleton without disassembling the cephalon (6, 7). Since other Phacopidid trilobites burrowed under the substrate before proceeding to molt, it is likely that Eldregreops also took this precautionary measure (7).
Trilobites at Seven Stars

Dipleura dekayi
As stated above, Dipleura is instantly recognizable because of its tuberculated preservation. It can grow up to at least 9 inches (23 cm) long, though specimens of these sizes are rare. This trilobite has a rounded, triangular, flattened cephalon, a wide, flat, triangular pygidium, and rounded segments with almost indistinguishable axial lobes.




Eldredgeops crassituberculata
Eldredgeops is exceedingly rare at Seven Stars, only known from one cephalon. It is more common at sites where there are fewer large, fast-swimming cephalopods; these at these sites, the spined Asteropygines seem to be more common. This pattern, seen in other sites too, may suggest that Eldredgeops shied away from large cephalopods, where Asteropygines such as Greenops and Bellacartwrightia were better suited because of their spines. Eldredgeops‘s color pattern has been fossilized; it had a pigmented exoskeleton with transparent spots to break up its outline.

Greenops boothi
Greenops boothi can be distinguished from Greenops grabaui because of its longer, less curved pyginal spines and slightly longer genal spines. Most common in the rocks to the left of the pullover, G. boothi has been known to grow to moderately large sizes at this site. The specimen at right is similar to others in that it preserves the color pattern of the live trilobite – the pyginal spines are darker than the axial lobe. See the life reconstruction of Greenops above to see how the rest of the trilobite was patterned.

Greenops grabaui
Greenops grabaui can be distinguished from Greenops boothi because of its shorter, more curves pyginal spines and slightly shorter genal spines. G. grabaui was otherwise very similar to G. boothi.
Works cited
(1) Wilson, K. A. (2014). Field guide to the Devonian fossils of New York (3rd ed.). Paleontological Research Institution.
(2) Gon, Sam, III. (2004-2024). A Guide to the Orders of Trilobites. trilobites.info.
This site no longer exits but was preserved on the Internet Archive, where it can be accessed as before.
(3) Fortey, Richard. (2010) Trilobite: Eyewitness to Evolution. Knopf Doubleday Publishing Group.
(4) Schoenemann, Brigitte & Clarkson, Euan & Bartels, C. & Südkamp, Wouter & Rössner, Gertrud & Ryck, Uwe. (2021). A 390 million-year-old hyper-compound eye in Devonian phacopid trilobites. Scientific Reports. 11. 10.1038/s41598-021-98740-z. https://www.researchgate.net/publication/354957278_A_390_million-year-old_hyper-compound_eye_in_Devonian_phacopid_trilobites
(5) Speyer, Stephen E. (1988). Biostratinomy and functional morphology of enrollment in two Middle Devonian trilobites. Lethaia, Vol. 21, pp. 121-138. Oslo. ISSN 0024-1164 www.thefossilforum.com/applications/core/interface/file/attachment.php?id=393425
This link leads to a HTTP file and may be a potential security risk.
(6) Zong, RW., Fan, RY. & Gong, YM. Seven 365-Million-Year-Old Trilobites Moulting within a Nautiloid Conch. Sci Rep 6, 34914 (2016). https://doi.org/10.1038/srep34914
(7) Drage HB, Daley AC. Recognising moulting behaviour in trilobites by examining morphology, development and preservation: Comment on Błażejowski et al. 2015. Bioessays. 2016 Oct;38(10):981-90. doi: 10.1002/bies.201600027. Epub 2016 Aug 22. PMID: 27545417; PMCID: PMC5042052. https://onlinelibrary.wiley.com/doi/full/10.1002/bies.201600027